
Seeing is believing: Visualization of DNA replication machinery in living cells
DNA replication is a fundamental process in every living organism. Precise regulation of genome duplication is absolutely critical for the fitness and survival of the organism as errors in DNA replication can lead to genome instability and the development of serious diseases such as cancer. Decades of beautiful work have provided new insights into the assembly, activation, and function of the replication fork, a basic unit of DNA replication also known as the replisome. However, the minichromosome maintenance 2-7 (MCM2-7) complex, the core of the replicative helicase, has puzzled cell biologists ever since the discovery of their essential role in DNA replication in the mid-90s. One of the striking facts about MCM biology ‘Why MCM2-7 proteins have never been visualized at DNA replication sites inside a cell?’ remained unanswered for several decades. Because this striking observation has been around for so long, scientists often refer to this puzzle as the MCM paradox. Studying the regulation of genome duplication in various cellular systems is the key to understanding the molecular basis of serious diseases of human health and designing new treatment strategies. Due to the inability to visualize the essential components of DNA replication machinery in the cellular environment, it has been difficult to pursue studies that advance the understanding of the regulation of genome duplication in cellular systems.
To break this deadlock, scientists from the Institute of Biophysics (IBP) of the Czech Academy of Sciences and the University of Copenhagen (UCPH) revisit the mysterious MCM paradox and their study published last week in Nature Communications solves the MCM paradox by genome editing enabling visualization of replication dynamics in living cells.
Dr. Hana Polasek-Sedlackova and her colleagues combined CRISPR-Cas9 genome editing of various replisome components with high-content imaging approach called QIBC (quantitative image-based cytometry) developed in Lukas lab. This technology allowed scientists to observe the initiation steps of the DNA replication process known as ‘origin licensing’ in the cellular environment. During origin licensing, MCM2-7 complexes are loaded on chromatin in an inactive state followed by its activation and the formation of the whole replisome machinery around the MCM scaffold at the beginning of the S phase. The IBP and UCPH scientists found that upon conversion of inactive MCM double hexamers to active replication forks, multiple replisome components block the antibody binding sites making the MCM scaffold of active replisome invisible in immunofluorescence-based experiments. They found that most of the MCM antibodies primarily detect the highly abundant inactive pool of MCMs, while CRISPR-Cas9 endogenous tagging of MCM subunits enables visualization of both inactive as well as the minor fraction of active MCMs in replication sites (see Figure 1).
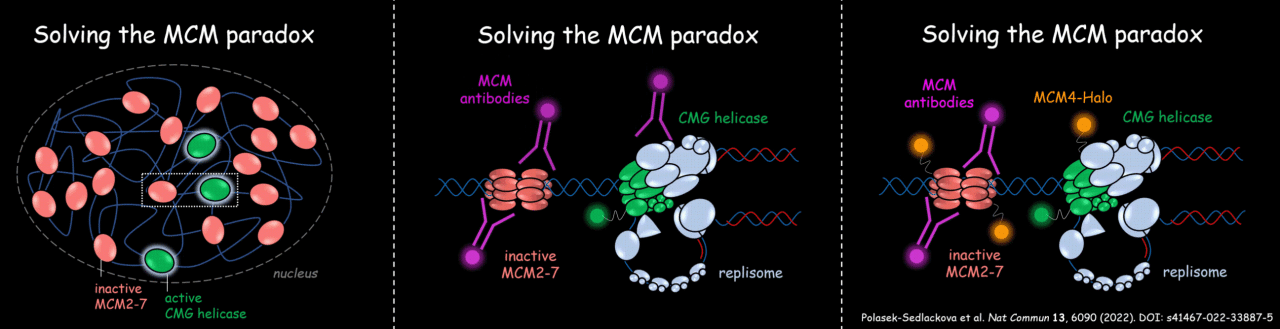
Figure 1: A graphical summary clarifying the MCM paradox.
In general, the findings of IBP and UCPH scientists uncovered potential limitations in studying large macromolecular biological complexes by immunofluorescence-based imaging approaches in the cellular environment. The multiple interacting partners of the protein of interest might shield antibody-binding sites, which can potentially complicate the interpretation of imaging data and studying basic molecular cellular processes in the cellular environment.
The work by IBP and UCPH scientists provides important validation of replisome architecture in physiological settings and opens new avenues for future investigation of replisome dynamics directly in living cells and understanding the molecular basis of serious human health diseases, such as cancer.
The complete story can be found here: https://www.nature.com/articles/s41467-022-33887-5
The work was funded by GACR junior star project (22-20303M), Novo Nordisk Foundation (NNF14CC0001), Danish National Research Foundation (DNRF115), and Carlsberg Foundation (CF21-0571).